


The fossil record is presented as the most decisive evidence for evolution. On closer reading, it is one of the strongest pieces of evidence against it. Life appears in the lowest fossil-bearing strata not in simple ancestral forms but in full diversity. The transitional forms Darwin’s theory requires are not there. The forms that ARE there have remained essentially unchanged for the periods of time the theory assigns them. The record is consistent with distinct creation followed by a single catastrophe; it is not consistent with the slow upward march of evolutionary mythology.
Charles Darwin understood the fossil record to be the most direct test of his theory. He wrote in On the Origin of Species, with characteristic honesty, that the geological record — if it failed to display the gradual succession of transitional forms his mechanism required — would constitute “the most obvious and gravest objection which can be urged against my theory.” A century and a half of fossil collection later, the test has been run. The result is not what he hoped for.
This article walks the fossil record in ten movements. Part I sets out what gradualism predicts. Part II examines the Cambrian explosion — the appearance of the major animal body plans, in full diversity, at the base of the fossil-bearing strata. Part III treats the continuing absence of the transitional forms the theory requires above the Cambrian. Part IV turns to the “living fossils” whose stasis over the conventional timescales contradicts the very mechanism that should have changed them. Part V looks at polystrate fossils — tree trunks passing vertically through multiple strata supposedly separated by millions of years — and what they testify to. Part VI examines the soft-tissue findings in dinosaur bones since 2005. Part VII reviews the most heavily promoted “transitional” claim of recent decades — the dinosaur-to-bird link — and the substantial scientific dissent against it. Part VIII reframes the spectrum of variation usually read as evolution. Part IX walks the mammalian fossil record and finds the opposite of evolution: a downward progression from giant ancestors to diminished modern descendants. Part X addresses the hominid story and the testimony of Christ on the question.
The companion article on Genesis vs Deep Time cross-examines the broader deep-time framework on which the evolutionary story depends, and the Flood carries the catastrophist case for the layered geology in which the fossils sit.
Part I — What gradualism predicts
The textbook account of the fossil record runs like this. In the oldest fossil-bearing strata, life appears in simple, single-celled forms. As the strata become younger, the organisms become more complex. Marine invertebrates give way to fishes; fishes give way to amphibians; amphibians to reptiles; reptiles to mammals and birds; and somewhere at the end of the sequence, primates and man. At every transition the chain is supposed to be filled with a sequence of intermediate forms, gradually changing one anatomical structure into another. The diagram in the front of every introductory biology textbook draws this as a branching tree, with the root in the simplest unicellular life and the tip in modern man.
That is the prediction the theory makes. There are two ways to test it. The first is to ask whether the lowest fossil-bearing strata actually show simple ancestors of the major animal groups. The second is to ask whether the higher strata show gradual transitional sequences between those groups. The honest answer to both questions, given the data the fossil record actually contains, is no.
Part II — The Cambrian explosion
The lowest fossil-bearing strata
The Precambrian strata, conventionally dated from the formation of the earth to about 541 million years ago, contain microbial mats, some single-celled organisms, and traces of soft-bodied multicellular life (the Ediacaran fauna being the principal example). What they do not contain is any ancestral form of the major animal phyla that appear in the strata above.
Then, at the base of the Cambrian, the fossil record transitions. In strata conventionally dated to a relatively brief window of approximately ten million years (the “Cambrian explosion” proper), the major animal body plans of the present day appear, essentially simultaneously, in fully developed form. Arthropods, molluscs, brachiopods, echinoderms, annelids, and chordates — the phylum to which vertebrates and ultimately humans belong — are all present at or near the base of the Cambrian. They appear without antecedents. They appear in their fully differentiated body plans.
“The Big Bang of animal evolution”
The honesty of the mainstream literature on this point is striking. Jeffrey S. Levinton, professor and chair of the Department of Ecology and Evolution at State University of New York at Stony Brook, summarised the situation in Scientific American in November 1992 under the title “The Big Bang of Animal Evolution”:
The Cambrian explosion was characterised by the sudden and roughly simultaneous appearance of many diverse animal forms almost 600 million years ago. No other period in the history of animal life can match this remarkable burst of evolutionary creativity.
The phrase is honest about the data. It is also a theological problem in disguise. “Sudden and roughly simultaneous appearance” of “many diverse animal forms” is the observation. The standard model has to explain it. It has not, in the thirty years since the article appeared, succeeded.
Our own phylum, present at the base
For most of the twentieth century, the standard textbook account held that the chordate phylum — to which all vertebrates, including humans, belong — was a late evolutionary arrival, appearing in the Ordovician period long after the Cambrian explosion. The 1990s discoveries from the Chengjiang fauna of Yunnan Province, China, overturned that account. The Chengjiang deposits, conventionally dated to roughly 525 million years ago (early Cambrian), contain not only well-preserved arthropods and other invertebrates but identifiable chordates — the small fish-like animal Myllokunmingia and the related Haikouichthys being the most striking examples.
The implication of this discovery, when it has been frankly stated by mainstream paleontologists, is decisive. The chordate phylum was present from the beginning. There is no Precambrian ancestor of the chordates. There is no sequence of intermediate forms leading up to them. They appear in the Cambrian, in fully formed body plans, along with every other major animal phylum on earth.
Gould’s rescue: punctuated equilibrium
The most serious attempt to accommodate the Cambrian explosion to the evolutionary framework was the theory of punctuated equilibrium proposed by Stephen Jay Gould and Niles Eldredge in 1972. Punctuated equilibrium holds that evolution proceeds in long periods of stasis interrupted by brief bursts of rapid change. The Cambrian “explosion” is, on this account, one such burst.
The rescue is honest about the data — it acknowledges that the transitional forms are not in the record — but it offers no mechanism by which large-scale anatomical innovation can occur in geologically brief windows. It postulates the rapid change to account for the absence of the slow change the data does not show. The result is a theory that remains consistent with the gaps in the record by declining to explain how those gaps could have been crossed at all. This is not, by any ordinary scientific standard, an improvement on the original theory. It is the original theory acknowledging that its principal predictions have not been confirmed.
Part III — The continuing absence of transitional forms
Darwin’s own difficulty
Darwin understood from the beginning that his theory required innumerable transitional forms, and that their conspicuous absence from the fossil record of his day was the most serious objection his theory had to answer. He wrote in On the Origin of Species:
Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely graduated organic chain; and this, perhaps, is the most obvious and gravest objection which can be urged against my theory. The explanation lies, as I believe, in the extreme imperfection of the geological record.
The defence was the “imperfection of the geological record”: the transitional forms had existed, but had simply not been preserved as fossils, or had been destroyed. The defence was reasonable in 1859. It rested on the hope that further excavation would supply the missing intermediates.
A century and a half of intensive fossil collection, across every continent and every formation, has not supplied them. The fossil record has been many times enlarged. Hundreds of thousands of new species have been catalogued. The systematic gaps Darwin pointed to, between the major groups of organisms, remain.
Gould’s admission
Stephen Jay Gould, no friend of creationism, was forthright about the data in his professional writings for paleontologists:
The extreme rarity of transitional forms in the fossil record persists as the trade secret of paleontology. The evolutionary trees that adorn our textbooks have data only at the tips and nodes of their branches; the rest is inference, however reasonable, not the evidence of fossils.
The phrase “trade secret of paleontology” is striking. The professionals in the field knew that the data did not match the textbook story. They taught the textbook story anyway. The data at the tips and nodes — the major groups themselves — were real. The lines that connected them on the diagrams in the textbooks were drawings, not fossils.
Where the gradualism should be visible
The strongest case for gradualism, if it could be made anywhere, would be made at the transitions between the major animal groups: invertebrates to fish, fish to amphibians, amphibians to reptiles, reptiles to mammals and birds. The textbook diagrams show each transition with confidence. The fossil record does not.
When a fish appears in the fossil record, it appears as a fish — fully fish, with fins, gills, scales, and a fish’s skeleton. When an amphibian appears, it appears as an amphibian, fully formed. When a bat appears, it appears as a bat, with the wing membrane already specialised for flight. When a whale appears, it appears as a whale. The systematic intermediate forms required by gradualism are not present. The phyla and the classes appear, like the body plans of the Cambrian, suddenly and fully formed, at their respective places in the column.
Part IV — The witness of the living fossils
If gradualism is the rule, then the organisms preserved in the deepest fossil-bearing strata should, by the time we encounter their living descendants today, have accumulated very substantial changes. The mechanism requires it. Random mutation acting on a population across hundreds of millions of years should produce enormous morphological divergence.
The data shows the opposite. A substantial number of modern organisms are essentially indistinguishable from their oldest fossil ancestors. The mainstream literature calls these “living fossils.” They are not anomalies. They span every kingdom of life and every conceivable habitat. A representative sample:
| Organism | Conventional dating | Living counterpart |
|---|---|---|
| Coelacanth | Lobe-finned fish. Declared extinct since the end of the Cretaceous, ~66 million years. | Caught alive off South Africa in 1938 by a fisherman, identified by Marjorie Courtenay-Latimer and J.L.B. Smith. Hundreds of specimens since. Anatomy essentially unchanged from the Cretaceous fossils. |
| Horseshoe crab | Marine arthropod. Conventionally dated to the Ordovician, ~450 million years. | Living populations on the Atlantic coast of North America and across East and South-East Asia. Identical in form to the Ordovician fossils. |
| Lingula brachiopod | Inarticulate brachiopod. Conventionally dated to the early Cambrian, ~520 million years — the base of the multicellular fossil record. | Living lingula in modern tidal flats from Japan to Australia. Microscopically indistinguishable from the Cambrian specimens. |
| Tuatara | Reptile of the order Rhynchocephalia. The order is conventionally dated to the Triassic, ~240 million years; thought extinct since the late Cretaceous. | Living tuatara on offshore islands of New Zealand. The order survives, in this one species, unchanged. |
| Ginkgo biloba | Gymnosperm tree. Conventionally dated to the Permian, ~270 million years. | Living ginkgoes are cultivated worldwide. The leaf pattern and reproductive structures match the Permian fossils. |
| Nautilus | Cephalopod with chambered shell. Conventionally dated to the Cambrian. | Living nautilus in the deep Pacific. Indistinguishable in form from its earliest fossil ancestors. |
The pattern continues across hundreds of taxa: cycad trees, dragonflies, certain sharks, lampreys, the Wollemi pine, the dawn redwood. Each is, on the conventional dating, supposed to have been subjected to hundreds of millions of years of evolutionary pressure. Each has remained essentially unchanged.
The mainstream rescue is “stabilising selection” — the suggestion that these organisms have found niches so stable that no significant evolutionary pressure has acted on them. The rescue has the same structural problem as the Gouldian rescue of the Cambrian explosion. It accounts for the absence of change by postulating the absence of the very pressure that should have driven it. It is consistent with the data only because it predicts no data. A theory which predicts both change (in the transitional gaps) and stasis (in the living fossils) and offers no principle to determine which to expect where, is a theory which predicts nothing in particular.
Part V — Polystrate trees and rapid burial
What the standard story requires
The standard reading of the layered sedimentary strata holds that they accumulated slowly, over hundreds of millions of years, with each layer representing a long period of geological time. The fossils within each layer are taken to represent the life of that period.
For a fossil to form, the organism in question must be buried before it decays. In modern environments, an unburied dead organism — a fish on the surface of the sea floor, a fallen tree in a forest — decomposes within months to a few years. It is not preserved. Fossilisation requires rapid burial under fine sediment, sufficient to exclude oxygen and the scavengers and microbes that would otherwise consume the organism, before decomposition advances.
The polystrate phenomenon
A polystrate fossil is one which passes vertically through multiple sedimentary strata representing, on the conventional dating, thousands or millions of years between deposition events. The most striking examples are upright fossilised tree trunks, often metres tall, passing through multiple coal seams and intervening shale or sandstone layers.
The classic site is the Joggins Fossil Cliffs on the Bay of Fundy in Nova Scotia — a UNESCO World Heritage Site. The cliffs preserve more than seventy coal seams interbedded with shale, and within those seams stand more than sixty polystrate lycopsid trees, some five and six metres tall, passing vertically through multiple strata. Identical polystrate trees occur at Yellowstone’s Specimen Ridge, in the Cumberland coalfields of Britain, in the Yellow Cat member of the Cedar Mountain Formation in Utah, and at dozens of other locations worldwide.
What the polystrate tree witnesses to
A polystrate tree cannot have stood upright through the supposedly long periods between successive strata. Above-ground decomposition rates would have toppled and decayed the trunk long before the next layer of sediment arrived. The trunk must have been buried — rapidly, completely, in a single depositional event covering its full length — or it could not have been preserved in the upright position the fossil shows.
Conventional geology now acknowledges this and treats polystrate fossils as evidence of localised catastrophic deposition events (volcanic ash falls, flash floods, deltaic surges). But every single polystrate tree, on every continent, requires its own local catastrophe on this reading — sometimes dozens at a single site. The Mt St Helens eruption of 1980, which buried thousands of trees in a few hours in flow deposits up to twenty-five feet thick, demonstrates the catastrophist mechanism. A single worldwide flood event is a more economical hypothesis than the dozens of independent local catastrophes the conventional reading requires. The fuller flood-geology case is in the companion article on the Flood.
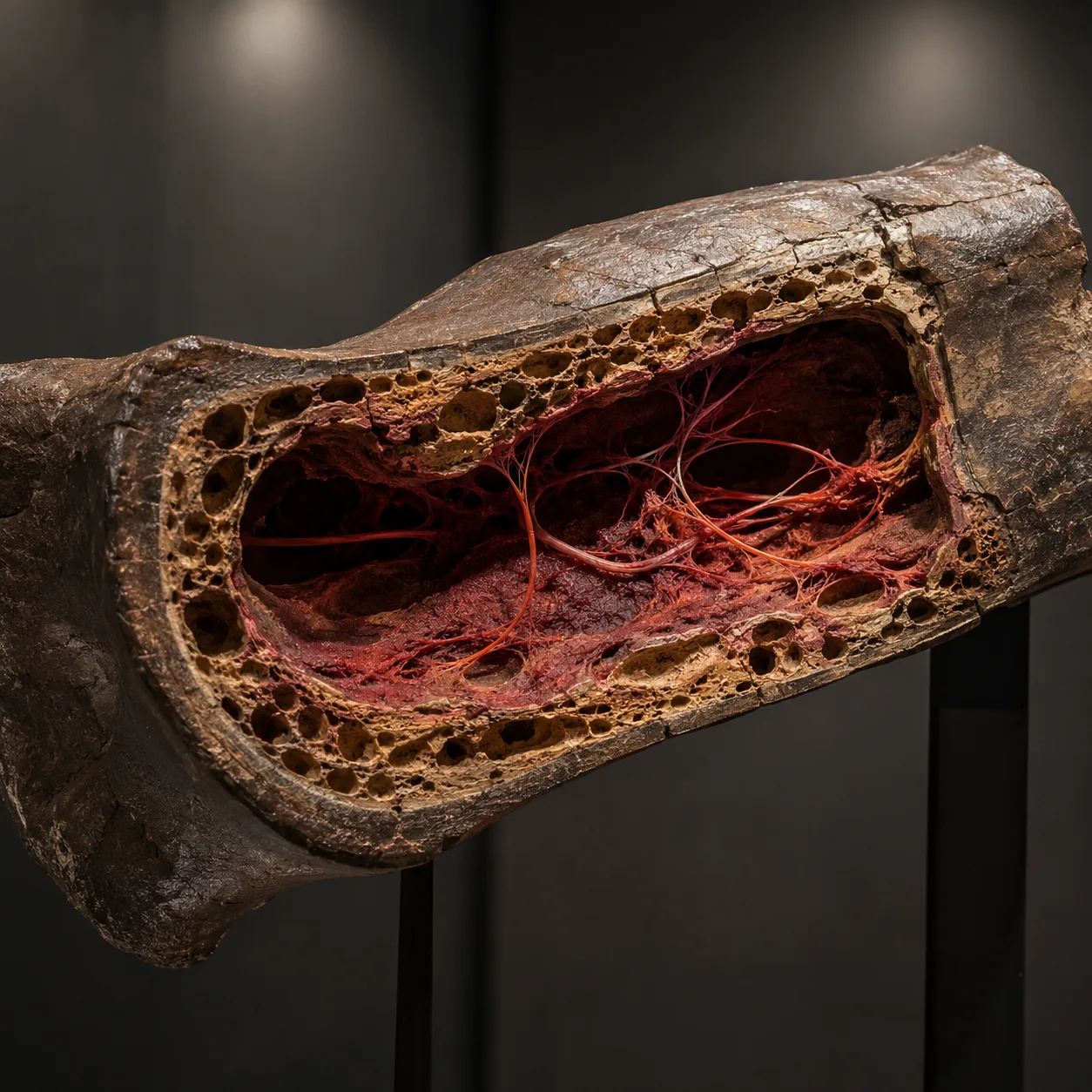
Part VI — Soft tissue in dinosaur bones
Schweitzer 2005
In March 2005, the palaeontologist Mary Schweitzer of North Carolina State University published a paper in the journal Science that has reverberated through her field ever since. Working on a femur fragment from a Tyrannosaurus rex specimen excavated from the Hell Creek Formation in Montana (the specimen is conventionally dated to approximately 68 million years before the present), Schweitzer dissolved the surrounding bone mineral with weak acid and recovered, from the inside of the bone:
- Soft, flexible tissue that stretched when pulled with tweezers.
- Branching blood vessels containing what appeared under microscopy to be intact red blood cells.
- Osteocytes (bone cells) preserving the characteristic stellate cell morphology.
- Fibrous matrix retaining the microscopic architecture of original collagen.
The finding has since been replicated in multiple subsequent papers and across multiple dinosaur specimens (a hadrosaur recovered later from Montana, a range of further Late Cretaceous and Jurassic specimens). In 2007 and again in 2017, mass-spectrometry sequencing has recovered actual amino-acid sequences from T. rex collagen protein — sequences that, on phylogenetic analysis, group most closely with modern chickens.
The decay-rate problem
The difficulty for the conventional dating is one of basic biochemistry. Soft tissue, intact cells, and identifiable protein sequences do not, by any laboratory model of organic decay, survive for sixty-eight million years. The protein collagen has a measured decomposition half-life that depends on temperature and burial environment, but under any plausible burial conditions the upper bound on detectable collagen survival is on the order of one to two million years — not sixty-eight. The data exceed the theoretical upper bound by a factor of thirty to fifty.
The mainstream response, advanced by Schweitzer herself, is that iron ions liberated from blood haemoglobin during decay provide a chemical preservation mechanism that vastly extends protein stability. The hypothesis is testable. Laboratory experiments with iron-saturated tissues have shown extended preservation — on the order of a few years extended into a few decades, not millions of years extended into tens of millions. The mechanism is real but does not, on the laboratory data, supply the three-to-five-orders-of-magnitude extension required to reconcile the find with the conventional dating.
The simpler reading
The simpler reading is that the bones are not 68 million years old. They are the remains of animals buried in a recent catastrophic event — recent enough that their soft tissue is still recoverable. On that reading, the soft-tissue findings are not anomalies requiring exotic preservation mechanisms. They are exactly what one would expect to find. The conventional date is the problem; the data is fine.
The same reading explains why fully articulated, three-dimensional dinosaur skeletons — rather than the scattered, partly-mineralised fragments slow-burial would produce — turn up repeatedly in flood-style depositional matrices around the world.
Part VII — The contested dinosaur-to-bird claim
The claim, and its public face
The claim that birds descended from theropod dinosaurs is the most heavily promoted “transitional” narrative of the last several decades. It is featured in nearly every children’s book on dinosaurs, in National Geographic specials, in museum dioramas, and in introductory biology textbooks. The standard presentation includes a series of supposedly intermediate fossils — Archaeopteryx, Sinosauropteryx, Microraptor, Anchiornis, and the briefly notorious Archaeoraptor, exposed as a chimera composite in 2000 — alleged to document the gradual transition from feathered theropod to flying bird.
The substantial dissent
What the public presentation does not generally acknowledge is the substantial body of professional dissent against the dinosaur-to-bird claim, mounted by specialists in avian palaeontology and physiology. Three voices in particular bear listening to:
| Source | Statement |
|---|---|
| Storrs L. Olson — Smithsonian National Museum of Natural History | Open letter to the President of the National Geographic Society, 1 November 1999, in response to the magazine's "Archaeoraptor" feature: "The idea of feathered dinosaurs and the theropod origin of birds is being actively promulgated by a cadre of zealous scientists acting in concert with certain editors at Nature and National Geographic… truth and careful scientific weighing of evidence have been among the first casualties in their program, which is now fast becoming one of the grander scientific hoaxes of our age — the paleontological equivalent of cold fusion." |
| Alan Feduccia — University of North Carolina | Quoted in The Origin and Evolution of Birds (Yale University Press, 1999): "The theropod-origin theory of birds, in my opinion, will be the greatest embarrassment of paleontology of the 20th century." Feduccia's case rests on the embryonic-thumb evidence (theropods have digits 1-2-3; birds have digits 2-3-4 in embryonic development) and the incompatibility of theropod respiratory anatomy with the avian flow-through lung. |
| John A. Ruben — Oregon State University | Published in Science (1997) on his analysis of Sinosauropteryx and Scipionyx specimens: the internal organs preserved show a bellows-style reptilian lung architecture, incompatible with the rigid through-flow lung required for avian respiration. The change from one architecture to the other cannot be accomplished by gradual modification; the intermediate stage would be a non-functional lung. |
The lung-architecture problem identified by Ruben is the most decisive of the three. Reptilian lungs (the lungs of theropod dinosaurs, on the standard reconstruction) work by inflation and deflation — air flows in, then back out, through the same structure. Avian lungs work by through-flow — air flows continuously in one direction through a network of rigid parabronchi, supplied by anterior and posterior air sacs. The two architectures are incompatible. No intermediate stage between them is functionally viable. A creature with a partly converted lung would not be able to respire at all. The transition cannot be made by the gradual modification the evolutionary mechanism requires.
The embryonic-thumb evidence
Feduccia’s embryonic-thumb argument is the second decisive piece of evidence. The five digits of the vertebrate hand are numbered 1 (thumb) to 5 (little finger). In the embryonic development of theropod dinosaurs, the digits that develop are 1, 2, and 3 — the thumb, index, and middle fingers — with 4 and 5 lost. In the embryonic development of birds, the digits that develop are 2, 3, and 4 — the index, middle, and ring fingers — with 1 and 5 lost. Theropods and birds have three-fingered hands, but the three fingers are not the same three fingers. The digits cannot be homologised. The hand of the bird is not developmentally descended from the hand of the theropod.
The mainstream response is the “frame shift” hypothesis — the suggestion that embryonic development was somehow shifted, in the course of evolution, so that the digits that develop look like 2-3-4 but are functionally homologous to 1-2-3. The hypothesis preserves the textbook conclusion at the cost of an unverifiable embryonic mechanism inserted ad hoc. As Feduccia put it, the theropod-origin theory of birds remains, in his view, the candidate for the greatest embarrassment of paleontology in the twentieth century.
Part VIII — Variety within kinds
What Darwin actually observed
The argument from the Galápagos finches that Darwin presented in On the Origin of Speciesand that has been retold in every introductory biology text since is, taken on its own merits, an entirely unobjectionable observation. The finches of the Galápagos are a single closely related group with substantial variation in beak shape and feeding habit between the islands. The beak shape variations correspond to local food sources. Selection pressure, acting on existing variation in beak shape, produces the local adaptations Darwin observed.
What the textbook account adds, and what the observation does not support, is the inference that this same mechanism, given enough time, will turn a finch into a non-finch — into a hawk, a whale, or a man. Variation within an existing kind is uncontroversial. Genesis describes God making the creatures “after their kind” (Gen 1:21, 24, 25) with the genetic capacity to vary widely within that kind. The dog breeds are the standard demonstration: the modern variation from Chihuahua to Saint Bernard, from Afghan hound to pug, is enormous, and it has all happened within the last several thousand years of selective breeding. None of it has produced anything other than a dog.
The thought experiment
A useful thought experiment will sharpen the distinction. Suppose a palaeontologist of the future, knowing nothing of modern dogs, dug up the skeletons of a Chihuahua, a fox terrier, a Labrador, a Great Dane, and a Saint Bernard, and was asked to construct an evolutionary tree from the bone evidence alone. He would, by ordinary palaeontological method, place the smallest skull at the base, the largest at the apex, and draw a series of intermediate forms in between. He would speak with confidence of evolutionary progression across the dog lineage. He would be wrong on every count. The animals lived contemporaneously. The variation was not evolutionary but selectional. The “tree” he constructed would be an artefact of the textbook narrative imposed on a body of fully co-existing variants of one underlying kind.
The same observation, applied across the fossil record, deflates a substantial fraction of the alleged “transitions.” The horse series is the textbook case. The supposed evolutionary lineage from the small Hyracotherium (formerly Eohippus) to the modern Equus arranges fossils of differing sizes and toe-counts in an apparently progressive series. The arrangement is made in the laboratory by the palaeontologist, not by the strata in which the fossils were found. Stratigraphically, the various forms overlap one another in time. Today, the modern horse breeds range in size from the Falabella miniature (about 30 inches tall) to the Shire horse (about 70 inches), all within the past few thousand years of selective breeding. The fossil “sequence” is consistent with variation within the horse kind, not with evolution between kinds.
Part IX — Mammals: devolution, not evolution
The age of giants
The fossil record of mammals presents one further difficulty for the evolutionary story. If evolution proceeds from small and simple toward large and complex — as Edward Drinker Cope’s much-cited “Cope’s rule” would have it — then the modern mammalian fauna should be, on average, larger and more diverse than the fauna of the Pleistocene that preceded it. The opposite is the case. The Pleistocene was the age of giants.
The list is long and well-attested. Mastodons and mammoths, twice the height of modern elephants. Glyptodons, armadillos the size of a Volkswagen. Giant ground sloths the height of a giraffe. Giant beavers the size of a black bear. Giant kangaroos and giant wombats in Australia. Cave bears half again the mass of modern grizzlies. Dire wolves. Sabre-toothed cats. Irish elk with antler spans of nearly four metres. Every group of mammals had its giants. Every group has lost them in the modern fauna.
The pattern as devolution
The pattern is the opposite of what the evolutionary story predicts. Modern mammals are not the apex of an upward progression. They are the diminished remnant of a fauna that, in the comparatively recent past, contained vastly more impressive specimens. The consistent direction of the change is downward — toward smaller body size, narrower distribution, less diversity, more extinction.
The straightforward biblical reading accounts for the pattern. The original creation was unfallen and unconstrained. The post-flood world was a degraded environment in which the genetic capacity for giantism, while still present, has been progressively lost as populations have bottlenecked, sub-speciated, and gone extinct. The companion article After Eden (forthcoming) treats the curse and its biological consequences in more detail.
The reframing of the dinosaurs
A parallel reframing applies to the dinosaurs. The textbook portrayal of Tyrannosaurus rex as a fearsome predator depends on the assumption that the animal’s teeth and stature must indicate carnivory. A closer look at T. rex anatomy tells a more complicated story. The forelimbs are tiny and could not have grasped struggling prey. The teeth, while long and pointed, are set in shallow sockets typical of bone-crushing scavengers rather than predators which must hold and tear. The locomotion of a six-tonne biped does not permit the sustained high-speed pursuit a major carnivore requires.
Modern animals with similar dentition include several fruit bats and the giant panda — whose impressive canines and crushing molars serve, in fact, to process tough plant matter. The Genesis account places all the original animals in a vegetarian condition (Gen 1:30) and Isaiah’s prophecy of the new earth (Isa 11:6–9) returns them to it. The fearsome teeth of T. rex may, on the biblical reading, have served originally for the same purpose modern panda teeth serve: the shredding of vegetation. The companion article After Eden takes the argument further.
Part X — The hominid story and the testimony of Christ
What the hominid fossils actually are
The textbook progression from Australopithecus through Homo habilis, Homo erectus, Homo heidelbergensis, and Homo neanderthalensis to modern Homo sapiens is the public face of human evolution. The implication conveyed by the diagrams — a series of progressively more human-looking creatures on a steady line — is that the transition from ape to man is documented in the fossil record.
The actual situation is more candidly described by the specialists. The australopithecines — A. afarensis (“Lucy”), A. africanus, A. robustus and the rest — are, on the published anatomical evidence, apes. Cranial capacity in the australopithecine range (375–550 cc) is well within the modern ape range. The published reconstructions of bipedal locomotion in A. afarensis rely heavily on the interpretation of the Laetoli footprints and on the contested reconstruction of the knee joint — which, in the case of the Lucy specimen, was recovered from a different stratum a considerable distance from the rest of the skeleton.
The Homo specimens, from habilis upward, fall within the wide range of normal human anatomical variation, with the neanderthals indistinguishable from modern humans except in robust build and slightly larger cranial capacity. Neanderthal DNA has now been sequenced from multiple specimens and is found to differ from modern human DNA by less than 1% on most measures. They are us.
What the textbook diagram presents as a smooth transition is, on examination, a body of fully ape fossils on one side, a body of fully human (with ordinary human variation) fossils on the other, and a set of contested intermediate cases that the specialists themselves disagree about. The gap is not bridged. It is photographed across.
The historical hoaxes
The history of the hominid “evidence” is itself instructive. Piltdown Man, accepted as a transitional form from 1912 to 1953, was revealed in 1953 to be a deliberate hoax assembled from a modern human skull and an orang-utan mandible. Nebraska Man, reconstructed in 1922 from a single tooth and pictured in the press as a full ape-man with wife and family, was identified in 1927 as the tooth of an extinct peccary — that is, a pig. Java Man (Pithecanthropus erectus) was assembled from a skullcap and a femur recovered fifty feet apart, the femur indistinguishable in fact from a modern human femur. Each hoax was, for decades, the centrepiece of the public case for human evolution. Each was eventually exposed by careful examination, often only after the next generation of students had been taught it as settled fact.
The testimony of Christ on the question
The question of human origins is not, in the end, a question on which the institute is content to rest its case on the contested palaeontology. The Lord Himself has spoken on it.
But from the beginning of the creation God made them male and female.
And he answered and said unto them, Have ye not read, that he which made them at the beginning made them male and female.
Christ’s phrase is “from the beginning of the creation.” He places the creation of the first man and the first woman at the beginning of the creation itself — not at the end of a billion-year evolutionary sequence. The reader who accepts Christ as the Son of God and the appointed teacher of His people is faced, on the question of human origins, with a choice of authority. The institute’s position rests on the testimony of Christ. The fossil record, on careful examination, does not contradict it.
Closing
The fossil record was offered by Darwin as the most decisive test of his theory. The test has been run. The intermediate forms his mechanism required have not been found. The forms that have been found appear suddenly, fully formed, and remain — where their lineages survive at all — essentially unchanged for the periods of time the theory assigns them. The mammalian fauna shows downward progression rather than upward. The most heavily promoted transition — dinosaurs to birds — collapses on the lung architecture and the embryonic digits. The hominid sequence resolves, on examination, into apes on one side and humans (with ordinary variation) on the other. The data does not support the textbook story.
The biblical reading accounts for the data simply. God created distinct kinds with genetic capacity for wide variation within each kind. A single catastrophic flood deposited the layered strata in which the fossils are found, fixing the patterns of mass burial, articulated skeletons, polystrate trees, marine fossils on mountain tops, and rapid sedimentation that the uniformitarian model has to invoke dozens of separate local catastrophes to explain. The post-flood environment was a diminished one, in which the original giantism has been progressively lost. The pattern is not evolution. It is variation, catastrophe, and devolution — in that order.
The witness of Scripture
And God created great whales, and every living creature that moveth, which the waters brought forth abundantly, after their kind, and every winged fowl after his kind: and God saw that it was good.
And God said, Let the earth bring forth the living creature after his kind, cattle, and creeping thing, and beast of the earth after his kind: and it was so. And God made the beast of the earth after his kind, and cattle after their kind, and every thing that creepeth upon the earth after his kind: and God saw that it was good.
But from the beginning of the creation God made them male and female.
For the invisible things of him from the creation of the world are clearly seen, being understood by the things that are made, even his eternal power and Godhead; so that they are without excuse.
But God giveth it a body as it hath pleased him, and to every seed his own body. All flesh is not the same flesh: but there is one kind of flesh of men, another flesh of beasts, another of fishes, and another of birds.
Through faith we understand that the worlds were framed by the word of God, so that things which are seen were not made of things which do appear.
Further Reading
- Walter J. Veith. Genesis Conflict, Lecture 103 (“Bones in Stones”) and Lecture 104 (“Where Mammals Reigned”). Amazing Discoveries. The principal source on which the survey in this article rests.
- Mary H. Schweitzer et al. “Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex.” Science 307: 1952–1955 (2005). The original soft-tissue paper.
- Alan Feduccia. The Origin and Evolution of Birds. Yale University Press, 1999. The principal scholarly critique of the dinosaur-to-bird theory.
- Storrs L. Olson. Open letter to the President of the National Geographic Society, 1 November 1999. The “Archaeoraptor” chimera-fossil incident and its aftermath.
- Jeffrey S. Levinton. “The Big Bang of Animal Evolution.” Scientific American, November 1992. The mainstream description of the Cambrian explosion.
- Stephen Jay Gould. “Evolution’s Erratic Pace.” Natural History 86 (May 1977). The “trade secret of paleontology” admission.
- Companion article: Genesis vs Deep Time — the broader deep-time framework on which the evolutionary story rests.
- Companion article: The Flood — the catastrophist account of the layered geology in which the fossils are preserved.
- Companion article: A Day to Be Remembered — the six literal days of creation week and the chronology built on them.
Foundational text
“And God made the beast of the earth after his kind, and cattle after their kind, and every thing that creepeth upon the earth after his kind: and God saw that it was good.”
— Genesis 1:25