


El registro fósil se presenta como la evidencia más decisiva a favor de la evolución. Leído con más cuidado, es una de las piezas de evidencia más fuertes en su contra. La vida aparece en los estratos fosilíferos más bajos no en formas ancestrales simples, sino en plena diversidad. Las formas transicionales que la teoría de Darwin requiere no están ahí. Las formas que SÍ están ahí han permanecido esencialmente sin cambios durante los lapsos de tiempo que la teoría les asigna. El registro es consistente con una creación distinta seguida de una sola catástrofe; no es consistente con la lenta marcha ascendente de la mitología evolutiva.
Charles Darwin entendía que el registro fósil era la prueba más directa de su teoría. Escribió en El origen de las especies, con su característica honestidad, que el registro geológico — si no llegaba a mostrar la sucesión gradual de formas transicionales que su mecanismo requería — constituiría “la objeción más obvia y grave que puede esgrimirse contra mi teoría.” Un siglo y medio de recolección de fósiles después, la prueba se ha realizado. El resultado no es el que él esperaba.
Este artículo recorre el registro fósil en diez movimientos. La Parte I expone lo que el gradualismo predice. La Parte II examina la explosión cámbrica — la aparición de los principales planes corporales animales, en plena diversidad, en la base de los estratos fosilíferos. La Parte III trata la ausencia continuada de las formas transicionales que la teoría requiere por encima del Cámbrico. La Parte IV se vuelve hacia los “fósiles vivientes” cuya estasis a lo largo de los lapsos convencionales contradice el mismísimo mecanismo que debió haberlos cambiado. La Parte V examina los fósiles polistratos — troncos de árbol que atraviesan verticalmente múltiples estratos supuestamente separados por millones de años — y de qué dan testimonio. La Parte VI examina los hallazgos de tejido blando en huesos de dinosaurio desde 2005. La Parte VII pasa revista a la afirmación “transicional” más intensamente promovida de las décadas recientes — el vínculo dinosaurio-ave — y al sustancial disenso científico en su contra. La Parte VIII reencuadra el espectro de variación que suele leerse como evolución. La Parte IX recorre el registro fósil de los mamíferos y halla lo opuesto de la evolución: una progresión descendente desde ancestros gigantes hasta descendientes modernos disminuidos. La Parte X aborda la historia de los homínidos y el testimonio de Cristo sobre la cuestión.
El artículo complementario sobre Génesis frente al tiempo profundo somete a contrainterrogatorio el marco más amplio del tiempo profundo del cual depende la historia evolutiva, y el Diluvio lleva el caso catastrofista de la geología estratificada en la que los fósiles yacen.
Parte I — Lo que el gradualismo predice
El relato de manual del registro fósil dice así. En los estratos fosilíferos más antiguos, la vida aparece en formas simples, unicelulares. A medida que los estratos se vuelven más jóvenes, los organismos se vuelven más complejos. Los invertebrados marinos ceden el paso a los peces; los peces ceden el paso a los anfibios; los anfibios a los reptiles; los reptiles a los mamíferos y las aves; y en algún punto al final de la secuencia, los primates y el hombre. En cada transición la cadena debe estar rellena de una secuencia de formas intermedias, transformando gradualmente una estructura anatómica en otra. El diagrama al frente de cada libro de texto introductorio de biología dibuja esto como un árbol ramificado, con la raíz en la vida unicelular más simple y la punta en el hombre moderno.
Esa es la predicción que la teoría hace. Hay dos maneras de ponerla a prueba. La primera es preguntar si los estratos fosilíferos más bajos realmente muestran ancestros simples de los principales grupos animales. La segunda es preguntar si los estratos superiores muestran secuencias transicionales graduales entre esos grupos. La respuesta honesta a ambas preguntas, dados los datos que el registro fósil realmente contiene, es no.
Parte II — La explosión cámbrica
Los estratos fosilíferos más bajos
Los estratos precámbricos, datados convencionalmente desde la formación de la tierra hasta hace unos 541 millones de años, contienen tapetes microbianos, algunos organismos unicelulares y rastros de vida multicelular de cuerpo blando (siendo la fauna ediacárica el ejemplo principal). Lo que no contienen es ninguna forma ancestral de los principales filos animales que aparecen en los estratos de arriba.
Entonces, en la base del Cámbrico, el registro fósil hace una transición. En estratos datados convencionalmente en una ventana relativamente breve de aproximadamente diez millones de años (la “explosión cámbrica” propiamente dicha), los principales planes corporales animales del presente aparecen, esencialmente de modo simultáneo, en forma plenamente desarrollada. Artrópodos, moluscos, braquiópodos, equinodermos, anélidos y cordados — el filo al que pertenecen los vertebrados y, en última instancia, los humanos — están todos presentes en la base del Cámbrico o cerca de ella. Aparecen sin antecedentes. Aparecen en sus planes corporales plenamente diferenciados.
“El big bang de la evolución animal”
La honestidad de la literatura corriente sobre este punto es llamativa. Jeffrey S. Levinton, profesor y director del Departamento de Ecología y Evolución de la State University of New York en Stony Brook, resumió la situación en Scientific American en noviembre de 1992 bajo el título “El big bang de la evolución animal”:
La explosión cámbrica se caracterizó por la aparición súbita y aproximadamente simultánea de muchas formas animales diversas hace casi 600 millones de años. Ningún otro período en la historia de la vida animal puede igualar este notable estallido de creatividad evolutiva.
La frase es honesta acerca de los datos. Es también un problema teológico disfrazado. La “aparición súbita y aproximadamente simultánea” de “muchas formas animales diversas” es la observación. El modelo estándar tiene que explicarla. No lo ha logrado, en los treinta años desde que apareció el artículo.
Nuestro propio filo, presente en la base
Durante la mayor parte del siglo XX, el relato estándar de manual sostenía que el filo de los cordados — al que pertenecen todos los vertebrados, incluyendo los humanos — fue una llegada evolutiva tardía, que apareció en el período Ordovícico mucho después de la explosión cámbrica. Los descubrimientos de la década de 1990 de la fauna de Chengjiang de la provincia de Yunnan, China, derribaron ese relato. Los depósitos de Chengjiang, datados convencionalmente en aproximadamente 525 millones de años (Cámbrico temprano), contienen no solo artrópodos y otros invertebrados bien preservados, sino cordados identificables — el pequeño animal con forma de pez Myllokunmingia y el relacionado Haikouichthys siendo los ejemplos más llamativos.
La implicación de este descubrimiento, cuando ha sido francamente declarada por los paleontólogos corrientes, es decisiva. El filo de los cordados estuvo presente desde el principio. No hay ningún ancestro precámbrico de los cordados. No hay ninguna secuencia de formas intermedias que conduzca hasta ellos. Aparecen en el Cámbrico, en planes corporales plenamente formados, junto con todos los demás filos animales principales de la tierra.
El rescate de Gould: el equilibrio puntuado
El intento más serio de acomodar la explosión cámbrica al marco evolutivo fue la teoría del equilibrio puntuado propuesta por Stephen Jay Gould y Niles Eldredge en 1972. El equilibrio puntuado sostiene que la evolución procede en largos períodos de estasis interrumpidos por breves estallidos de cambio rápido. La “explosión” cámbrica es, según este relato, uno de tales estallidos.
El rescate es honesto acerca de los datos — reconoce que las formas transicionales no están en el registro — pero no ofrece ningún mecanismo por el cual la innovación anatómica a gran escala pueda ocurrir en ventanas geológicamente breves. Postula el cambio rápido para dar cuenta de la ausencia del cambio lento que los datos no muestran. El resultado es una teoría que permanece consistente con las lagunas del registro al negarse a explicar cómo esas lagunas pudieron cruzarse en absoluto. Esto no es, según ningún estándar científico ordinario, una mejora sobre la teoría original. Es la teoría original reconociendo que sus principales predicciones no han sido confirmadas.
Parte III — La ausencia continuada de formas transicionales
La dificultad del propio Darwin
Darwin entendió desde el principio que su teoría requería innumerables formas transicionales, y que su conspicua ausencia del registro fósil de su época era la objeción más seria que su teoría tenía que responder. Escribió en El origen de las especies:
¿Por qué, entonces, no está cada formación geológica y cada estrato lleno de tales eslabones intermedios? La geología ciertamente no revela ninguna cadena orgánica tan finamente graduada; y esto, quizá, es la objeción más obvia y grave que puede esgrimirse contra mi teoría. La explicación reside, según creo, en la extrema imperfección del registro geológico.
La defensa fue la “imperfección del registro geológico”: las formas transicionales habían existido, pero simplemente no se habían preservado como fósiles, o habían sido destruidas. La defensa era razonable en 1859. Descansaba en la esperanza de que la excavación ulterior supliría los intermedios faltantes.
Un siglo y medio de intensa recolección de fósiles, a través de cada continente y cada formación, no los ha suplido. El registro fósil ha sido ampliado muchas veces. Cientos de miles de nuevas especies han sido catalogadas. Las lagunas sistemáticas que Darwin señaló, entre los principales grupos de organismos, permanecen.
La admisión de Gould
Stephen Jay Gould, nada amigo del creacionismo, fue franco acerca de los datos en sus escritos profesionales para paleontólogos:
La extrema rareza de las formas transicionales en el registro fósil persiste como el secreto comercial de la paleontología. Los árboles evolutivos que adornan nuestros libros de texto tienen datos solo en las puntas y los nodos de sus ramas; el resto es inferencia, por razonable que sea, no la evidencia de los fósiles.
La frase “secreto comercial de la paleontología” es llamativa. Los profesionales del campo sabían que los datos no concordaban con la historia de manual. Enseñaban la historia de manual de todos modos. Los datos en las puntas y los nodos — los grupos principales mismos — eran reales. Las líneas que los conectaban en los diagramas de los libros de texto eran dibujos, no fósiles.
Dónde debería verse el gradualismo
El caso más fuerte a favor del gradualismo, si pudiera hacerse en algún lugar, se haría en las transiciones entre los principales grupos animales: invertebrados a peces, peces a anfibios, anfibios a reptiles, reptiles a mamíferos y aves. Los diagramas de manual muestran cada transición con confianza. El registro fósil no.
Cuando un pez aparece en el registro fósil, aparece como un pez — plenamente pez, con aletas, branquias, escamas y el esqueleto de un pez. Cuando un anfibio aparece, aparece como un anfibio, plenamente formado. Cuando un murciélago aparece, aparece como un murciélago, con la membrana del ala ya especializada para el vuelo. Cuando una ballena aparece, aparece como una ballena. Las formas intermedias sistemáticas requeridas por el gradualismo no están presentes. Los filos y las clases aparecen, como los planes corporales del Cámbrico, de modo súbito y plenamente formados, en sus respectivos lugares de la columna.
Parte IV — El testimonio de los fósiles vivientes
Si el gradualismo es la regla, entonces los organismos preservados en los estratos fosilíferos más profundos deberían, para cuando encontremos a sus descendientes vivientes hoy, haber acumulado cambios muy sustanciales. El mecanismo lo requiere. La mutación aleatoria actuando sobre una población a lo largo de cientos de millones de años debería producir una enorme divergencia morfológica.
Los datos muestran lo opuesto. Un número sustancial de organismos modernos son esencialmente indistinguibles de sus más antiguos ancestros fósiles. La literatura corriente los llama “fósiles vivientes”. No son anomalías. Abarcan cada reino de la vida y cada hábitat concebible. Una muestra representativa:
| Organismo | Datación convencional | Contraparte viviente |
|---|---|---|
| Celacanto | Pez de aletas lobuladas. Declarado extinto desde el final del Cretácico, ~66 millones de años. | Capturado vivo frente a Sudáfrica en 1938 por un pescador, identificado por Marjorie Courtenay-Latimer y J.L.B. Smith. Cientos de especímenes desde entonces. Anatomía esencialmente sin cambios respecto de los fósiles cretácicos. |
| Cangrejo herradura | Artrópodo marino. Datado convencionalmente en el Ordovícico, ~450 millones de años. | Poblaciones vivientes en la costa atlántica de Norteamérica y por todo el Este y Sudeste de Asia. Idéntico en forma a los fósiles ordovícicos. |
| Braquiópodo Lingula | Braquiópodo inarticulado. Datado convencionalmente en el Cámbrico temprano, ~520 millones de años — la base del registro fósil multicelular. | Lingula viviente en las llanuras mareales modernas desde Japón hasta Australia. Microscópicamente indistinguible de los especímenes cámbricos. |
| Tuátara | Reptil del orden Rhynchocephalia. El orden se data convencionalmente en el Triásico, ~240 millones de años; se creía extinto desde el Cretácico tardío. | Tuátara viviente en islas costeras de Nueva Zelanda. El orden sobrevive, en esta única especie, sin cambios. |
| Ginkgo biloba | Árbol gimnosperma. Datado convencionalmente en el Pérmico, ~270 millones de años. | Los ginkgos vivientes se cultivan en todo el mundo. El patrón de las hojas y las estructuras reproductivas concuerdan con los fósiles pérmicos. |
| Nautilo | Cefalópodo de concha tabicada. Datado convencionalmente en el Cámbrico. | Nautilo viviente en el Pacífico profundo. Indistinguible en forma de sus más tempranos ancestros fósiles. |
El patrón continúa a través de cientos de taxones: las cícadas, las libélulas, ciertos tiburones, las lampreas, el pino de Wollemi, la secuoya del alba. Cada uno, según la datación convencional, se supone que ha estado sometido a cientos de millones de años de presión evolutiva. Cada uno ha permanecido esencialmente sin cambios.
El rescate corriente es la “selección estabilizadora” — la sugerencia de que estos organismos han hallado nichos tan estables que ninguna presión evolutiva significativa ha actuado sobre ellos. El rescate tiene el mismo problema estructural que el rescate gouldiano de la explosión cámbrica. Da cuenta de la ausencia de cambio postulando la ausencia de la mismísima presión que debió haberlo impulsado. Es consistente con los datos solo porque no predice ningún dato. Una teoría que predice tanto cambio (en las lagunas transicionales) como estasis (en los fósiles vivientes) y no ofrece ningún principio para determinar cuál esperar dónde, es una teoría que no predice nada en particular.
Parte V — Árboles polistratos y enterramiento rápido
Lo que la historia estándar requiere
La lectura estándar de los estratos sedimentarios estratificados sostiene que se acumularon lentamente, a lo largo de cientos de millones de años, representando cada capa un largo período de tiempo geológico. Los fósiles dentro de cada capa se toman como representativos de la vida de ese período.
Para que un fósil se forme, el organismo en cuestión debe ser enterrado antes de descomponerse. En los ambientes modernos, un organismo muerto sin enterrar — un pez en la superficie del fondo marino, un árbol caído en un bosque — se descompone en cuestión de meses a unos pocos años. No se preserva. La fosilización requiere un enterramiento rápido bajo sedimento fino, suficiente para excluir el oxígeno y los carroñeros y microbios que de otro modo consumirían el organismo, antes de que avance la descomposición.
El fenómeno polistrato
Un fósil polistrato es uno que atraviesa verticalmente múltiples estratos sedimentarios que representan, según la datación convencional, miles o millones de años entre eventos de deposición. Los ejemplos más llamativos son troncos de árbol fosilizados en posición vertical, a menudo de varios metros de altura, que atraviesan múltiples vetas de carbón y las capas intermedias de esquisto o arenisca.
El sitio clásico son los Acantilados Fósiles de Joggins en la Bahía de Fundy, en Nueva Escocia — un Sitio de Patrimonio Mundial de la UNESCO. Los acantilados preservan más de setenta vetas de carbón intercaladas con esquisto, y dentro de esas vetas se yerguen más de sesenta árboles licópsidos polistratos, algunos de cinco y seis metros de altura, que atraviesan verticalmente múltiples estratos. Árboles polistratos idénticos se hallan en Specimen Ridge de Yellowstone, en los yacimientos de carbón de Cumberland en Gran Bretaña, en el miembro Yellow Cat de la Formación Cedar Mountain en Utah, y en docenas de otras localidades en todo el mundo.
De qué da testimonio el árbol polistrato
Un árbol polistrato no pudo haber permanecido en posición vertical a través de los supuestamente largos períodos entre estratos sucesivos. Las tasas de descomposición sobre el suelo habrían derribado y descompuesto el tronco mucho antes de que llegara la siguiente capa de sedimento. El tronco debió haber sido enterrado — rápidamente, por completo, en un solo evento de deposición que cubrió toda su longitud — o no podría haberse preservado en la posición vertical que el fósil muestra.
La geología convencional reconoce ahora esto y trata los fósiles polistratos como evidencia de eventos de deposición catastrófica localizados (caídas de ceniza volcánica, inundaciones repentinas, oleadas deltaicas). Pero cada árbol polistrato individual, en cada continente, requiere su propia catástrofe local según esta lectura — a veces docenas en un solo sitio. La erupción del Monte Santa Helena de 1980, que enterró miles de árboles en unas pocas horas en depósitos de flujo de hasta siete metros y medio de espesor, demuestra el mecanismo catastrofista. Un solo evento de diluvio mundial es una hipótesis más económica que las docenas de catástrofes locales independientes que la lectura convencional requiere. El caso más completo de la geología del diluvio está en el artículo complementario sobre el Diluvio.
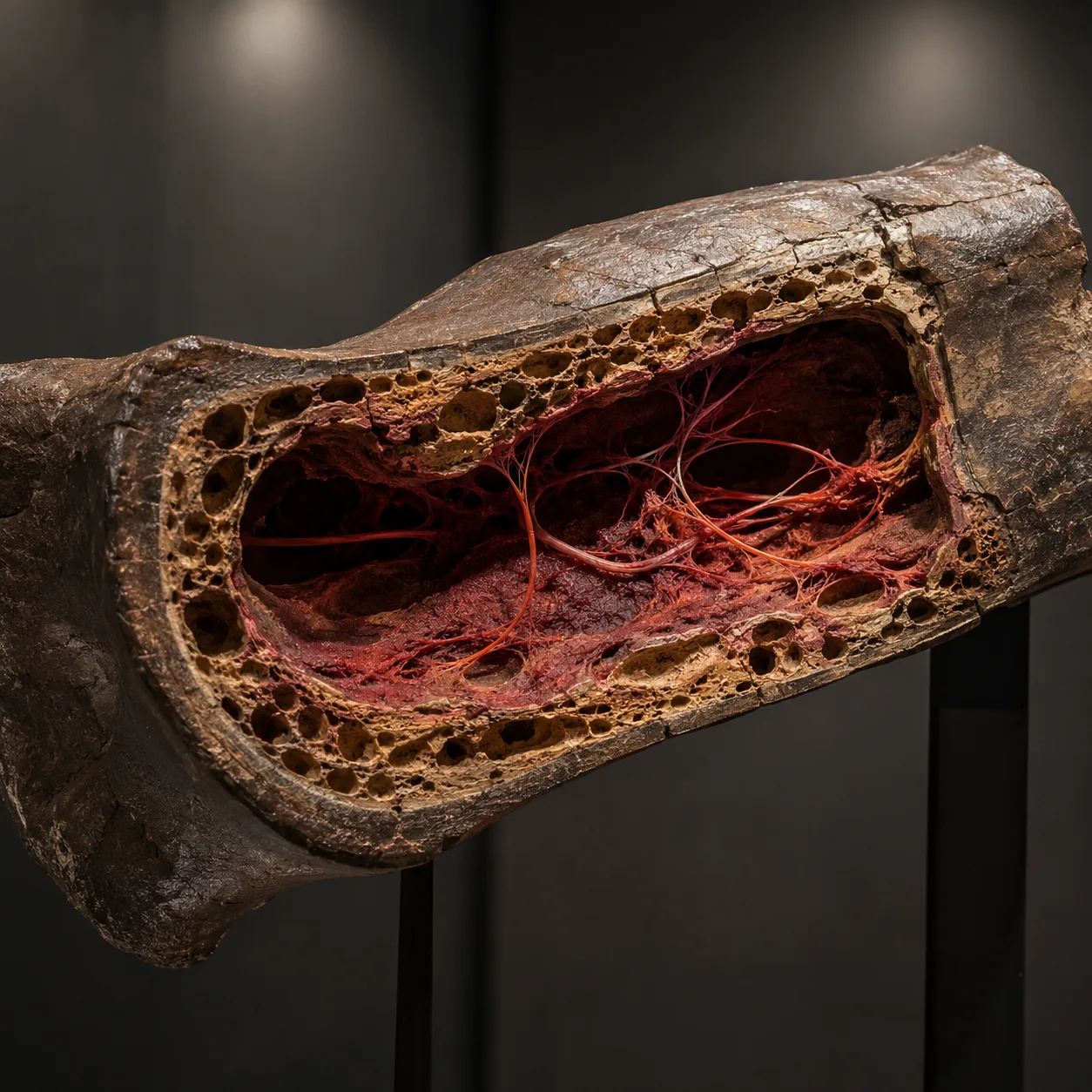
Parte VI — Tejido blando en huesos de dinosaurio
Schweitzer 2005
En marzo de 2005, la paleontóloga Mary Schweitzer de la North Carolina State University publicó un artículo en la revista Science que ha resonado en su campo desde entonces. Trabajando sobre un fragmento de fémur de un espécimen de Tyrannosaurus rex excavado de la Formación Hell Creek en Montana (el espécimen se data convencionalmente en aproximadamente 68 millones de años antes del presente), Schweitzer disolvió el mineral óseo circundante con ácido débil y recuperó, del interior del hueso:
- Tejido blando y flexible que se estiraba al ser jalado con pinzas.
- Vasos sanguíneos ramificados que contenían lo que parecían bajo el microscopio glóbulos rojos intactos.
- Osteocitos (células óseas) que preservaban la característica morfología celular estrellada.
- Matriz fibrosa que retenía la arquitectura microscópica del colágeno original.
El hallazgo ha sido replicado desde entonces en múltiples artículos posteriores y a través de múltiples especímenes de dinosaurio (un hadrosaurio recuperado más tarde en Montana, una gama de especímenes adicionales del Cretácico tardío y del Jurásico). En 2007 y de nuevo en 2017, la secuenciación por espectrometría de masas ha recuperado secuencias reales de aminoácidos de la proteína de colágeno de T. rex — secuencias que, en el análisis filogenético, se agrupan más estrechamente con los pollos modernos.
El problema de la tasa de descomposición
La dificultad para la datación convencional es de bioquímica básica. El tejido blando, las células intactas y las secuencias proteicas identificables no sobreviven, según ningún modelo de laboratorio de descomposición orgánica, durante sesenta y ocho millones de años. La proteína colágeno tiene una vida media de descomposición medida que depende de la temperatura y del ambiente de enterramiento, pero bajo cualquier condición plausible de enterramiento el límite superior de supervivencia detectable de colágeno es del orden de uno a dos millones de años — no sesenta y ocho. Los datos exceden el límite superior teórico por un factor de treinta a cincuenta.
La respuesta corriente, propuesta por la propia Schweitzer, es que los iones de hierro liberados de la hemoglobina de la sangre durante la descomposición proveen un mecanismo de preservación química que extiende vastamente la estabilidad proteica. La hipótesis es comprobable. Los experimentos de laboratorio con tejidos saturados de hierro han mostrado una preservación extendida — del orden de unos pocos años extendidos a unas pocas décadas, no de millones de años extendidos a decenas de millones. El mecanismo es real, pero no provee, según los datos de laboratorio, la extensión de tres a cinco órdenes de magnitud requerida para reconciliar el hallazgo con la datación convencional.
La lectura más simple
La lectura más simple es que los huesos no tienen 68 millones de años. Son los restos de animales enterrados en un evento catastrófico reciente — lo bastante reciente como para que su tejido blando todavía sea recuperable. Según esa lectura, los hallazgos de tejido blando no son anomalías que requieran mecanismos exóticos de preservación. Son exactamente lo que cabría esperar hallar. La fecha convencional es el problema; los datos están bien.
La misma lectura explica por qué esqueletos de dinosaurio plenamente articulados y tridimensionales — en lugar de los fragmentos dispersos y parcialmente mineralizados que produciría un enterramiento lento — aparecen repetidamente en matrices de deposición de estilo diluvial por todo el mundo.
Parte VII — La disputada afirmación dinosaurio-ave
La afirmación y su cara pública
La afirmación de que las aves descienden de dinosaurios terópodos es la narrativa “transicional” más intensamente promovida de las últimas décadas. Aparece en casi todo libro infantil sobre dinosaurios, en especiales de National Geographic, en dioramas de museo y en libros de texto introductorios de biología. La presentación estándar incluye una serie de fósiles supuestamente intermedios — Archaeopteryx, Sinosauropteryx, Microraptor, Anchiornis y el brevemente célebre Archaeoraptor, expuesto como una quimera compuesta en 2000 — supuestamente para documentar la transición gradual de terópodo emplumado a ave voladora.
El sustancial disenso
Lo que la presentación pública no suele reconocer es el sustancial cuerpo de disenso profesional contra la afirmación dinosaurio-ave, planteado por especialistas en paleontología y fisiología aviar. Tres voces en particular merecen ser escuchadas:
| Fuente | Declaración |
|---|---|
| Storrs L. Olson — Museo Nacional de Historia Natural del Smithsonian | Carta abierta al Presidente de la National Geographic Society, 1 de noviembre de 1999, en respuesta al reportaje de la revista sobre el «Archaeoraptor»: la idea de los dinosaurios emplumados y del origen terópodo de las aves está siendo promovida activamente por un grupo de científicos celosos que actúan de común acuerdo con ciertos editores de Nature y National Geographic; la verdad y el pesaje científico cuidadoso de la evidencia han sido las primeras víctimas de su programa, que se está convirtiendo rápidamente en uno de los mayores fraudes científicos de nuestra era — el equivalente paleontológico de la fusión fría. |
| Alan Feduccia — Universidad de Carolina del Norte | Citado en The Origin and Evolution of Birds (Yale University Press, 1999): la teoría del origen terópodo de las aves, en su opinión, será la mayor vergüenza de la paleontología del siglo XX. El argumento de Feduccia descansa en la evidencia del pulgar embrionario (los terópodos tienen los dígitos 1-2-3; las aves tienen los dígitos 2-3-4 en el desarrollo embrionario) y en la incompatibilidad de la anatomía respiratoria terópoda con el pulmón aviar de flujo continuo. |
| John A. Ruben — Universidad Estatal de Oregón | Publicado en Science (1997) sobre su análisis de los especímenes de Sinosauropteryx y Scipionyx: los órganos internos preservados muestran una arquitectura pulmonar reptiliana de tipo fuelle, incompatible con el pulmón rígido de flujo continuo requerido para la respiración aviar. El cambio de una arquitectura a la otra no puede lograrse por modificación gradual; la etapa intermedia sería un pulmón no funcional. |
El problema de la arquitectura pulmonar identificado por Ruben es el más decisivo de los tres. Los pulmones reptilianos (los pulmones de los dinosaurios terópodos, según la reconstrucción estándar) funcionan por inflación y deflación — el aire entra, luego sale, a través de la misma estructura. Los pulmones aviares funcionan por flujo continuo — el aire fluye continuamente en una sola dirección a través de una red de parabronquios rígidos, abastecidos por sacos aéreos anteriores y posteriores. Las dos arquitecturas son incompatibles. Ninguna etapa intermedia entre ellas es funcionalmente viable. Una criatura con un pulmón parcialmente convertido no podría respirar en absoluto. La transición no puede hacerse por la modificación gradual que el mecanismo evolutivo requiere.
La evidencia del pulgar embrionario
El argumento del pulgar embrionario de Feduccia es la segunda pieza decisiva de evidencia. Los cinco dígitos de la mano de los vertebrados se numeran del 1 (pulgar) al 5 (meñique). En el desarrollo embrionario de los dinosaurios terópodos, los dígitos que se desarrollan son el 1, el 2 y el 3 — el pulgar, el índice y el dedo medio — perdiéndose el 4 y el 5. En el desarrollo embrionario de las aves, los dígitos que se desarrollan son el 2, el 3 y el 4 — el índice, el medio y el anular — perdiéndose el 1 y el 5. Los terópodos y las aves tienen manos de tres dedos, pero los tres dedos no son los mismos tres dedos. Los dígitos no pueden homologarse. La mano del ave no desciende, desde el punto de vista del desarrollo, de la mano del terópodo.
La respuesta corriente es la hipótesis del “desplazamiento de marco” — la sugerencia de que el desarrollo embrionario fue de algún modo desplazado, en el curso de la evolución, de manera que los dígitos que se desarrollan parecen 2-3-4 pero son funcionalmente homólogos al 1-2-3. La hipótesis preserva la conclusión de manual a costa de un mecanismo embrionario inverificable insertado ad hoc. Como lo planteó Feduccia, la teoría del origen terópodo de las aves sigue siendo, a su juicio, la candidata a la mayor vergüenza de la paleontología del siglo XX.
Parte VIII — Variedad dentro de los tipos
Lo que Darwin realmente observó
El argumento de los pinzones de las Galápagos que Darwin presentó en El origen de las especies y que se ha vuelto a contar en cada texto introductorio de biología desde entonces es, tomado en sus propios términos, una observación enteramente inobjetable. Los pinzones de las Galápagos son un único grupo estrechamente emparentado con variación sustancial en la forma del pico y el hábito alimenticio entre las islas. Las variaciones en la forma del pico corresponden a las fuentes de alimento locales. La presión de selección, actuando sobre la variación existente en la forma del pico, produce las adaptaciones locales que Darwin observó.
Lo que el relato de manual añade, y que la observación no respalda, es la inferencia de que este mismo mecanismo, dado suficiente tiempo, convertirá un pinzón en un no-pinzón — en un halcón, una ballena o un hombre. La variación dentro de un tipo existente es incontrovertible. El Génesis describe a Dios haciendo a las criaturas “según su especie” (Gén 1:21, 24, 25) con la capacidad genética para variar ampliamente dentro de ese tipo. Las razas de perros son la demostración estándar: la variación moderna desde el chihuahua hasta el san bernardo, desde el galgo afgano hasta el carlino, es enorme, y toda ella ha ocurrido en los últimos varios miles de años de cría selectiva. Nada de ello ha producido otra cosa que un perro.
El experimento mental
Un útil experimento mental afilará la distinción. Supóngase que un paleontólogo del futuro, sin saber nada de los perros modernos, desenterrara los esqueletos de un chihuahua, un fox terrier, un labrador, un gran danés y un san bernardo, y se le pidiera construir un árbol evolutivo solo a partir de la evidencia ósea. Él, por el método paleontológico ordinario, colocaría el cráneo más pequeño en la base, el más grande en el ápice, y dibujaría una serie de formas intermedias en medio. Hablaría con confianza de una progresión evolutiva a través del linaje canino. Se equivocaría en todos los puntos. Los animales vivieron de modo contemporáneo. La variación no fue evolutiva sino selectiva. El “árbol” que construyó sería un artefacto de la narrativa de manual impuesta sobre un cuerpo de variantes plenamente coexistentes de un solo tipo subyacente.
La misma observación, aplicada a todo el registro fósil, desinfla una fracción sustancial de las supuestas “transiciones”. La serie del caballo es el caso de manual. El supuesto linaje evolutivo desde el pequeño Hyracotherium (antes Eohippus) hasta el Equus moderno dispone fósiles de diferentes tamaños y números de dedos en una serie aparentemente progresiva. La disposición la hace en el laboratorio el paleontólogo, no los estratos en los que se hallaron los fósiles. Estratigráficamente, las diversas formas se solapan unas a otras en el tiempo. Hoy, las razas modernas de caballos varían en tamaño desde el miniatura Falabella (de unos 76 centímetros de altura) hasta el caballo Shire (de unos 178 centímetros), todo dentro de los últimos pocos miles de años de cría selectiva. La “secuencia” fósil es consistente con la variación dentro del tipo caballo, no con la evolución entre tipos.
Parte IX — Mamíferos: devolución, no evolución
La era de los gigantes
El registro fósil de los mamíferos presenta una dificultad más para la historia evolutiva. Si la evolución procede de lo pequeño y simple hacia lo grande y complejo — como tendría la muy citada “regla de Cope” de Edward Drinker Cope — entonces la fauna mamífera moderna debería ser, en promedio, mayor y más diversa que la fauna del Pleistoceno que la precedió. Es lo contrario lo que ocurre. El Pleistoceno fue la era de los gigantes.
La lista es larga y bien atestiguada. Mastodontes y mamuts, del doble de la altura de los elefantes modernos. Gliptodontes, armadillos del tamaño de un Volkswagen. Perezosos terrestres gigantes de la altura de una jirafa. Castores gigantes del tamaño de un oso negro. Canguros gigantes y wombats gigantes en Australia. Osos de las cavernas con la mitad más de masa que los osos pardos modernos. Lobos gigantes. Felinos dientes de sable. Alces irlandeses con cornamentas de casi cuatro metros de envergadura. Cada grupo de mamíferos tuvo sus gigantes. Cada grupo los ha perdido en la fauna moderna.
El patrón como devolución
El patrón es lo opuesto de lo que la historia evolutiva predice. Los mamíferos modernos no son el ápice de una progresión ascendente. Son el remanente disminuido de una fauna que, en el pasado comparativamente reciente, contenía especímenes mucho más impresionantes. La dirección consistente del cambio es descendente — hacia un tamaño corporal menor, una distribución más estrecha, menos diversidad, más extinción.
La lectura bíblica directa da cuenta del patrón. La creación original era no caída y sin restricciones. El mundo posdiluviano fue un ambiente degradado en el que la capacidad genética para el gigantismo, aunque todavía presente, se ha perdido progresivamente a medida que las poblaciones han sufrido cuellos de botella, se han subespeciado y se han extinguido. El artículo complementario Después del Edén (próximamente) trata la maldición y sus consecuencias biológicas con más detalle.
El reencuadre de los dinosaurios
Un reencuadre paralelo se aplica a los dinosaurios. El retrato de manual del Tyrannosaurus rex como un temible depredador depende de la suposición de que los dientes y la estatura del animal deben indicar carnivoría. Una mirada más atenta a la anatomía de T. rex cuenta una historia más complicada. Las extremidades anteriores son diminutas y no podrían haber aferrado una presa que forcejeara. Los dientes, aunque largos y puntiagudos, están asentados en alvéolos superficiales típicos de los carroñeros trituradores de huesos más que de los depredadores que deben sostener y desgarrar. La locomoción de un bípedo de seis toneladas no permite la persecución sostenida a alta velocidad que un gran carnívoro requiere.
Entre los animales modernos con una dentición similar se incluyen varios murciélagos frugívoros y el oso panda gigante — cuyos impresionantes caninos y molares trituradores sirven, de hecho, para procesar materia vegetal dura. El relato del Génesis ubica a todos los animales originales en una condición vegetariana (Gén 1:30) y la profecía de Isaías sobre la nueva tierra (Isa 11:6–9) los devuelve a ella. Los temibles dientes de T. rex pueden, en la lectura bíblica, haber servido originalmente para el mismo propósito que sirven los dientes del panda moderno: la trituración de la vegetación. El artículo complementario Después del Edén lleva el argumento más lejos.
Parte X — La historia de los homínidos y el testimonio de Cristo
Qué son realmente los fósiles de homínidos
La progresión de manual desde Australopithecus a través de Homo habilis, Homo erectus, Homo heidelbergensis y Homo neanderthalensis hasta el moderno Homo sapiens es la cara pública de la evolución humana. La implicación transmitida por los diagramas — una serie de criaturas progresivamente más parecidas a lo humano sobre una línea continua — es que la transición de simio a hombre está documentada en el registro fósil.
La situación real la describen con más franqueza los especialistas. Los australopitecos — A. afarensis (“Lucy”), A. africanus, A. robustus y el resto — son, según la evidencia anatómica publicada, simios. La capacidad craneal en el rango de los australopitecos (375–550 cc) está bien dentro del rango de los simios modernos. Las reconstrucciones publicadas de la locomoción bípeda en A. afarensis se apoyan fuertemente en la interpretación de las huellas de Laetoli y en la disputada reconstrucción de la articulación de la rodilla — que, en el caso del espécimen de Lucy, se recuperó de un estrato diferente a una distancia considerable del resto del esqueleto.
Los especímenes de Homo, desde habilis hacia arriba, caen dentro del amplio rango de la variación anatómica humana normal, con los neandertales indistinguibles de los humanos modernos salvo en la constitución robusta y la capacidad craneal ligeramente mayor. El ADN neandertal se ha secuenciado ahora de múltiples especímenes y se halla que difiere del ADN humano moderno en menos del 1% en la mayoría de las mediciones. Son nosotros.
Lo que el diagrama de manual presenta como una transición fluida es, examinado de cerca, un cuerpo de fósiles plenamente simios de un lado, un cuerpo de fósiles plenamente humanos (con variación humana ordinaria) del otro, y un conjunto de casos intermedios disputados sobre los que los propios especialistas discrepan. La brecha no está salvada. Está fotografiada por encima.
Los fraudes históricos
La historia de la “evidencia” de los homínidos es ella misma instructiva. El Hombre de Piltdown, aceptado como forma transicional desde 1912 hasta 1953, se reveló en 1953 como un fraude deliberado ensamblado a partir de un cráneo humano moderno y una mandíbula de orangután. El Hombre de Nebraska, reconstruido en 1922 a partir de un solo diente y retratado en la prensa como un hombre-simio completo con esposa y familia, se identificó en 1927 como el diente de un pecarí extinto — es decir, un cerdo. El Hombre de Java (Pithecanthropus erectus) se ensambló a partir de una calota y un fémur recuperados a quince metros de distancia, siendo el fémur de hecho indistinguible de un fémur humano moderno. Cada fraude fue, durante décadas, la pieza central del caso público a favor de la evolución humana. Cada uno fue eventualmente expuesto por un examen cuidadoso, a menudo solo después de que a la siguiente generación de estudiantes se le hubiera enseñado como hecho establecido.
El testimonio de Cristo sobre la cuestión
La cuestión de los orígenes humanos no es, al final, una cuestión sobre la cual el instituto se contente con descansar su caso en la paleontología disputada. El Señor mismo ha hablado sobre ella.
Pero al principio de la creación, varón y hembra los hizo Dios.
Y él respondiendo, les dijo: ¿No habéis leído que el que los hizo al principio, varón y hembra los hizo?
La frase de Cristo es “al principio de la creación”. Él ubica la creación del primer hombre y de la primera mujer en el principio de la creación misma — no al final de una secuencia evolutiva de mil millones de años. El lector que acepta a Cristo como el Hijo de Dios y el maestro designado de Su pueblo se enfrenta, en la cuestión de los orígenes humanos, a una elección de autoridad. La posición del instituto descansa en el testimonio de Cristo. El registro fósil, examinado con cuidado, no lo contradice.
Conclusión
El registro fósil fue ofrecido por Darwin como la prueba más decisiva de su teoría. La prueba se ha realizado. Las formas intermedias que su mecanismo requería no se han hallado. Las formas que sí se han hallado aparecen de modo súbito, plenamente formadas, y permanecen — donde sus linajes sobreviven en absoluto — esencialmente sin cambios durante los lapsos de tiempo que la teoría les asigna. La fauna mamífera muestra una progresión descendente en lugar de ascendente. La transición más intensamente promovida — de dinosaurios a aves — se desmorona ante la arquitectura pulmonar y los dígitos embrionarios. La secuencia de los homínidos se resuelve, examinada, en simios de un lado y humanos (con variación ordinaria) del otro. Los datos no respaldan la historia de manual.
La lectura bíblica da cuenta de los datos de modo simple. Dios creó tipos distintos con capacidad genética para amplia variación dentro de cada tipo. Un solo diluvio catastrófico depositó los estratos estratificados en los que se hallan los fósiles, fijando los patrones de enterramiento masivo, esqueletos articulados, árboles polistratos, fósiles marinos en las cimas de las montañas, y sedimentación rápida que el modelo uniformista tiene que invocar docenas de catástrofes locales separadas para explicar. El ambiente posdiluviano fue uno disminuido, en el que el gigantismo original se ha perdido progresivamente. El patrón no es evolución. Es variación, catástrofe y devolución — en ese orden.
El testimonio de la Escritura
Y crió Dios las grandes ballenas, y toda cosa viva que anda arrastrando, que las aguas produjeron según su género, y toda ave alada según su especie: y vió Dios que era bueno.
Y dijo Dios: Produzca la tierra seres vivientes según su género, bestias y serpientes y animales de la tierra según su especie: y fué así. É hizo Dios animales de la tierra según su género, y ganado según su género, y todo animal que anda arrastrando sobre la tierra según su especie: y vió Dios que era bueno.
Pero al principio de la creación, varón y hembra los hizo Dios.
Porque las cosas invisibles de él, su eterna potencia y divinidad, se echan de ver desde la creación del mundo, siendo entendidas por las cosas que son hechas; de modo que son inexcusables.
Mas Dios le da el cuerpo como quiso, y á cada simiente su propio cuerpo. Toda carne no es la misma carne; mas una carne ciertamente es la de los hombres, y otra carne la de los animales, y otra la de los peces, y otra la de las aves.
Por la fe entendemos haber sido compuestos los siglos por la palabra de Dios, siendo hecho lo que se ve, de lo que no se veía.
Citas originales
Esta página es una recomposición en español del artículo original en inglés; los versículos bíblicos se citan de la RV1909. Las palabras de los testigos humanos citados arriba (científicos, revistas, cartas) se ofrecieron recompuestas en español; se reproducen abajo en su lengua de origen, el inglés. Los versículos bíblicos se excluyen de esta caja.
The Cambrian explosion was characterised by the sudden and roughly simultaneous appearance of many diverse animal forms almost 600 million years ago. No other period in the history of animal life can match this remarkable burst of evolutionary creativity.
Jeffrey S. Levinton — Scientific American, noviembre de 1992 · original en inglés
Why then is not every geological formation and every stratum full of such intermediate links? Geology assuredly does not reveal any such finely graduated organic chain; and this, perhaps, is the most obvious and gravest objection which can be urged against my theory. The explanation lies, as I believe, in the extreme imperfection of the geological record.
Charles Darwin — On the Origin of Species, ch. X · original en inglés
The extreme rarity of transitional forms in the fossil record persists as the trade secret of paleontology. The evolutionary trees that adorn our textbooks have data only at the tips and nodes of their branches; the rest is inference, however reasonable, not the evidence of fossils.
Stephen Jay Gould — Natural History, May 1977 · original en inglés
The idea of feathered dinosaurs and the theropod origin of birds is being actively promulgated by a cadre of zealous scientists acting in concert with certain editors at Nature and National Geographic… truth and careful scientific weighing of evidence have been among the first casualties in their program, which is now fast becoming one of the grander scientific hoaxes of our age — the paleontological equivalent of cold fusion.
Storrs L. Olson — open letter to the President of the National Geographic Society, 1 November 1999 · original en inglés
The theropod-origin theory of birds, in my opinion, will be the greatest embarrassment of paleontology of the 20th century.
Alan Feduccia — The Origin and Evolution of Birds (Yale University Press, 1999) · original en inglés
Fuentes
- Walter J. Veith. Genesis Conflict, Lecture 103 (“Bones in Stones”) y Lecture 104 (“Where Mammals Reigned”). Amazing Discoveries. La fuente principal sobre la que descansa el recorrido de este artículo.
- Mary H. Schweitzer et al. “Soft-Tissue Vessels and Cellular Preservation in Tyrannosaurus rex.” Science 307: 1952–1955 (2005). El artículo original sobre el tejido blando.
- Alan Feduccia. The Origin and Evolution of Birds. Yale University Press, 1999. La principal crítica académica de la teoría dinosaurio-ave.
- Storrs L. Olson. Carta abierta al Presidente de la National Geographic Society, 1 de noviembre de 1999. El incidente del fósil-quimera “Archaeoraptor” y sus secuelas.
- Jeffrey S. Levinton. “The Big Bang of Animal Evolution.” Scientific American, noviembre de 1992. La descripción corriente de la explosión cámbrica.
- Stephen Jay Gould. “Evolution’s Erratic Pace.” Natural History 86 (mayo de 1977). La admisión del “secreto comercial de la paleontología”.
- Artículo complementario: Génesis frente al tiempo profundo — el marco más amplio del tiempo profundo sobre el que descansa la historia evolutiva.
- Artículo complementario: El Diluvio — el relato catastrofista de la geología estratificada en la que se preservan los fósiles.
- Artículo complementario: Un día para recordar — los seis días literales de la semana de la creación y la cronología construida sobre ellos.
Texto fundacional
“É hizo Dios animales de la tierra según su género, y ganado según su género, y todo animal que anda arrastrando sobre la tierra según su especie: y vió Dios que era bueno.”
— Génesis 1:25 (RV1909)